Celleskjebnebestemmelse: "Integrert kompleksitet: Argumentet for intelligent design i cellulære kommunikasjonssystemer"
Av Otangelo Grasso -oversatt herfra.
 Kompleksiteten i cellulære koder og nødvendigheten av krysskommunikasjon i flercellede organismer: bevis for intelligent design
Kompleksiteten i cellulære koder og nødvendigheten av krysskommunikasjon i flercellede organismer: bevis for intelligent design
Konseptet med krysskommunikasjon mellom forskjellige cellulære koder antyder et høyt nivå av koordinering og formål i utformingen av biologiske systemer. De intrikate interaksjonene og kommunikasjonen mellom disse kodene indikerer et sofistikert og integrert design som lar celler svare på miljømessige signaler, opprettholde homeostase og utføre funksjonene sine effektivt. Kompleksiteten og spesifisiteten til disse kodene, og deres evne til å samhandle med hverandre, kan ikke forklares tilstrekkelig med en trinnvis, gradvis og ikke-styrt evolusjonsprosess. Eksemplene på krysskommuniasjon mellom de karakteriserte kodene fremhever hvordan forskjellige cellulære prosesser er sammenflettet, og endringer i en kode kan ha kaskaderende effekter på andre. For at disse komplekse reguleringssystemene skal fungere sammenhengende, må alle komponenter være på plass samtidig og samarbeide. Sannsynligheten for alle disse intrikate mekanismene som utvikler seg uavhengig gjennom tilfeldige mutasjoner og naturlig seleksjon er ekstremt lav, gitt den nøyaktige koordinasjonen som kreves for cellulær funksjon. For at flercellularitet skulle dukke opp og fungere effektivt, måtte flere nøkkelcellekoder være i full drift helt fra begynnelsen.
Bilde 1. Hensikten med kommunikasjon
De 31 genetiske kodene: Den genetiske koden er det universelle settet med regler som oversetter sekvensen av nukleotider i DNA og RNA til sekvensen av aminosyrer i proteiner. Proteiner er avgjørende for alle cellulære prosesser, og flercellede organismer er avhengige av et stort utvalg av proteiner for å utføre forskjellige funksjoner. Celleadhesjon er prosessen som holder celler sammen, noe som muliggjør dannelse av vev og organer. Uten celleadhesjon ville celler ikke være i stand til å organisere og samarbeide, og flercellede strukturer ville ikke være mulig. Denne koden styrer hvordan celler skiller og spesialiserer seg i forskjellige celletyper under utviklingen. Celleskjebnebestemmelse er viktig for dannelse av distinkte vev og organer med spesifikke funksjoner. Cellepolaritet refererer til den romlige organisasjonen og asymmetri i celler. Riktig cellepolaritet er avgjørende for celleposisjonering i vev og hjelper celler til å fungere kollektivt med å bygge komplekse strukturer. Kromatin er komplekset av DNA og proteiner som utgjør kromosomer. Kromatinkoden kontrollerer genuttrykk og regulerer cellulær differensiering, slik at celler kan ta på seg spesifikke funksjoner. Hox-genene spiller en betydelig rolle i å spesifisere kroppssegmentidentitet under utvikling, og sikrer riktig regionalisering av kroppen og dannelsen av kroppsstrukturer langs den fremre-posterior aksen.
Signaltransduksjon lar celler motta og svare på eksterne signaler. Det er viktig for å koordinere cellulære responser og atferd i flercellede organismer. Transkripsjonsfaktorer regulerer ekspresjonen av spesifikke gener, dirigerer cellulær differensiering og dannelse av forskjellige vev og celletyper. RNA -molekyler spiller forskjellige roller i cellulære prosesser, inkludert genregulering, katalyse og strukturell støtte. Disse funksjonene er avgjørende for flercellede organismer. DNA-metylering er en epigenetisk modifisering som påvirker  genuttrykk og celledifferensiering. Det hjelper med å etablere celleidentiteter og er essensiell for vevsspesifikke funksjoner i flercellede organismer. Disse kodene var essensielle helt fra begynnelsen av multicellularitet, fordi de ga den nødvendige koordinering og regulering for celler til å samarbeide, differensiere til spesialiserte celletyper og feste seg til hverandre, for å danne vev og etablere organiserte kroppsstrukturer. Uten disse kodene fullt operasjonelle ville ikke flercellede organismer ha vært i stand til å utvikle seg og fungere som sammenhengende og komplekse enheter.
genuttrykk og celledifferensiering. Det hjelper med å etablere celleidentiteter og er essensiell for vevsspesifikke funksjoner i flercellede organismer. Disse kodene var essensielle helt fra begynnelsen av multicellularitet, fordi de ga den nødvendige koordinering og regulering for celler til å samarbeide, differensiere til spesialiserte celletyper og feste seg til hverandre, for å danne vev og etablere organiserte kroppsstrukturer. Uten disse kodene fullt operasjonelle ville ikke flercellede organismer ha vært i stand til å utvikle seg og fungere som sammenhengende og komplekse enheter.
Bilde 2. Metylering er en måte å nedarve ervervede egenskaper
Koder som krysskommuniserer med andre koder
For å fungere ordentlig og muliggjøre kompleksiteten i flercellede organismer, trenger de fleste av disse kodene å kunne krysskommunisere med andre koder. Krysskommunikasjon og interaksjon mellom disse kodene muliggjør koordinert regulering av cellulære prosesser, vevsdannelse og generell kroppsarkitektur. Her er en sammentrekning av hvilke koder som trenger å krysskommunisere med andre:
Celleadhesjonskoden: samhandler med den EkstraCellulære Matrise (ECM) -koden og andre signalveier for celle-til-celleadhesjon og vevsdannelse.
Cellsskjebnebestemmelses-kode: Krysser med signaloverføringskoden og transkripsjonsfaktorkode for å bestemme celleskjebnen basert på miljøsignaler.
Cellpolaritetskoden: Krever interaksjon med cytoskjelett-koden og andre signalveier for å regulere cellulær organisering.
Kromatinkoden: interagerer med transkripsjons-faktorkoden og RNA -koden for å regulere genuttrykk og cellulær differensiering.
Hox-koden: samhandler med forskjellige transkripsjonsfaktorer og signalveier for å spesifisere kroppssegmentidentitet og koordinere kroppsstrukturdannelse.
Signaltransduksjonskoden: Krysskommunisere med andre koder, for eksempel transkripsjonsfaktorkode og cellesyklus sjekkpunktkode, for koordinerte cellulære responser.
Transkripsjonsfaktorkoden: samhandler med kromatinkoden, RNA -koden og andre for å regulere genuttrykk og cellulær differensiering.
RNA -koden: samhandler med ribosomal kode, RNA -redigeringskode og andre for genregulering og strukturell støtte.
DNA-metyleringskoden: Krysser med kromatinkoden og transkripsjonsfaktorkoden for å påvirke genuttrykk og celledifferensiering.
Disse kodene fungerer sammen for å muliggjøre celledifferensiering, vevsdannelse, genregulering og den  generelle flercellede organisasjonen. Deres interaksjoner og koordinering er avgjørende for utvikling og funksjon av komplekse flercellede organismer.
generelle flercellede organisasjonen. Deres interaksjoner og koordinering er avgjørende for utvikling og funksjon av komplekse flercellede organismer.
Bilde 3. Grasso konkluderer om genregulerende nettverk
Kompleksiteten og spesifisiteten til de cellulære kodene, deres evne og deres nødvendighet til å krysskommunisere med hverandre antyder et høyt nivå av koordinering og formål i utformingen av biologiske systemer. Disse kodene er intrikat sammenflettet, og endringer i en kode kan ha kaskaderende effekter på andre, noe som indikerer en sofistikert og integrert design som lar celler svare på miljøvennlige signaler og utføre funksjonene sine effektivt. Fremveksten av disse kodene på en trinnvis, evolusjonær måte ville utgjøre betydelige utfordringer. For at disse komplekse reguleringssystemene skal fungere sammenhengende, må alle komponenter være på plass samtidig og samarbeide. Det er lite sannsynlig at disse intrikate mekanismene kunne ha utviklet seg uavhengig gjennom tilfeldige mutasjoner og naturlig seleksjon fordi den koordinerte funksjonen av flere koder ville kreve et ekstraordinært antall spesifikke mutasjoner for å oppstå samtidig. Videre representerer kodene i seg selv svært spesifikke og informasjonsrike systemer. De involverer komplekse nettverk av interaksjoner mellom molekyler, signalveier og genregulerende elementer. For å fungere ordentlig, må disse systemene være fint innstilt og presise. Sannsynligheten for en slik presisjon som fremtredende trinn for trinn gjennom gradvise, ikke-styrte prosesser er ekstremt lav, i det umulige riket. I tillegg krever etablering av multicellularitet og utvikling av kompleks organismal arkitektur flere koder for å samarbeide på en koordinert måte. Den vellykkede integrasjonen av disse kodene er avgjørende for celledifferensiering, vevsdannelse og den generelle funksjonen til flercellede organismer. Det er usannsynlig å forvente at disse kodene uavhengig kunne ha utviklet seg og koordinert sine funksjoner over tid, da det ville kreve et astronomisk antall heldige hendelser. Tilstedeværelsen av disse fullt operasjonelle kodene helt fra begynnelsen av flercellularitet antyder bevisst planlegging og formål i utformingen av biologiske systemer. Evnen til disse kodene til å kryss-kommunisere og samhandle sømløst støtter ideen om en intelligent designer som orkestrerer de intrikate prosessene, som ligger til grunn for kompleksiteten i flercellulært liv.
Denne listen inkluderer 103 forskjellige biologiske cellepigenetiske koder. Det dekker et bredt spekter av cellulære prosesser og funksjoner, og fremhever kompleksiteten og mangfoldet av epigenetisk regulering i celler.
Tips: Opprett et nytt vindu med denne musikken, mens du kikker over lista.
1. De 31 genetiske kodene
2. Acetyleringskoden
3. De akustiske kodene
4. Vedhengsskoden
5. Acyleringskoden
6. Antioksidantkoden
7. Apoptosekoden
8. Den autokrine signalkoden
9. Autofagi -koden
10. Den bioelektriske koden
11. Biofotonkoden
12. Kalsiumkoden
13. Celleadhesjonskoden
14. Cellskjebne bestemmelseskode
15. Cellemigrasjonskoden
16. Cellepolaritets koden
17. Chaperonkoden
18. Kromatinkoden
19. Den kromosomale avtrykkskoden
20. Cilia -koden
21. Koden for sirkulært motiv (ribosom)
22. Cytoskjelettkoden
23. Koaktivatoren/corepressor/epigenetisk kode
24. Koden for menneskelig språk
25. Romkoden
26. Den skjulte koden i den genetiske koden
27. DNA-skadesvarekoden
28. DNA-metyleringskoden
29. Differensieringskoden
30. Domenesubstratspesifisitetskoden for ikke -ribosomale peptidsyntetaser (NRPs)
31. Endocytosekoden
32. Epidermal Vekst Faktor(EVF) -koden
33. Den epitranscriptomiske koden
34. Feilkorrigeringskoden
35. Den ekstracellulære matrisen (ECM) -koden
36. Den genomiske koden
37. Den genomiske reguleringskoden
38. G-ProteinKoblet Reseptor (GPKR) kode
39. Glykomi kode
40. Glykosyleringskoden
41. Hedgehog -signalkoden
42. Heterokromatinkoden
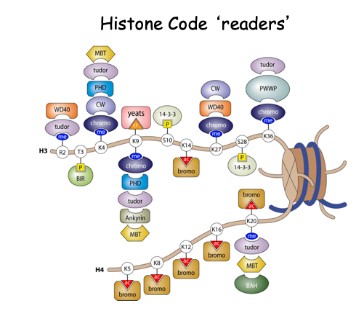 43. Histonvariantkoden -se eks. på Bilde 4. Histon-kode avlesere
43. Histonvariantkoden -se eks. på Bilde 4. Histon-kode avlesere
44. Hox-koden
45. Immunresponskoden eller språket
46. Inositol fosfatkode
47. Laminkoden
48. Metabolsk signalkode
49. Metyleringskoden
50. Mikrobiomkoden
51. NitrogenOksid (NO) signalkode
52. N-GLYCAN-koden
53. Den ikke-ribosomale koden
54. Nukleosom koden
55. Næringssensings koden
56. Myelinkoden
57. Den nevronale aktivitetsavhengige genuttrykkskoden
58. Den neuronale pigghastighetskoden
59. Den ikke-ribosomale koden
60. Nukleosomkoden
61. Atomsignalkoden
62. Olfaktorisk kode
63. Operonkoden
64. Fosforyleringskoden
65. Fosforyleringsavhengig proteininteraksjonskode
66. Fosfolipidkoden
67. Den post-translasjonelle modifikasjonskoden for transkripsjonsfaktorer
68. RNA -koden
69. Ribosomal kode
70. Ribosvitsj -koden
71. Proteinfoldingskoden
72. Protein Sekretorisk kode
73. Redox -koden
74. Retinsyresignalkoden
75. Ribonukleinsyremodifiserings-koden (RNA -modifiseringskode)
..
76. RNA -redigeringskoden
77. RNA -interferens (RNAi) -koden
78. Serotoninkoden
79. Splitekodene
80. Signaltransduksjonskoden
81. Signalintegrasjonskodene
82. Sukkerkoden
83. Den synaptiske limkoden
84. Stamcellekoden
85. Sumoylerings-koden
86. Talin -koden
87. Toll-Lignende Reseptor (TLR) kode
88. Transkripsjonsfaktor-koden
89. Den transkripsjonelle cis-regulatoriske koden
90. Transkripsjonell reguleringskode
91. Den transkripsjonelle cis-regulatoriske koden
92. Tubulin -koden
 93. Ubiquitin-koden
93. Ubiquitin-koden
94. Tumorsuppressor-koden
95. UBLIQUITIN-lignende modifikator (UBL) kode
96. Wnt -signalkoden
97. Kalsiumsignalkoden
09. Cellesyklus sjekkpunktkode
99. Cellecellekommunikasjons-koden
100. DNA-reparasjonskoden
101. Hormonreseptor-koden
102. Næringsstransport-koden
103. Fotosyntesekoden
Bilde 5. Annerledes virkning ved epigenetisk-'tilkoblinger'
Mange av de over 100 epigenetiske kodene krysskommuniserer med hverandre på grunn av sammenkoblingen av cellulære prosesser. Krysskommunikasjon refererer til kommunikasjonen og samspillet mellom forskjellige signalveier og reguleringsmekanismer i cellen.
Acetyleringskoden og metyleringskoden: Disse kodene kan samhandle i sammenheng med epigenetisk regulering, der acetylering og metylering av histoner kan modulere genuttrykk.
Autofagi-koden og næringssenseringskoden: Autofagi er regulert som respons på næringstilgjengelighet og cellulært energinivå, noe som indikerer krysskommunikasjon mellom disse kodene i cellulær metabolisme.
DNA-skaderesponskoden og DNA-reparasjonskoden: DNA-skaderesponsen aktiverer DNA-reparasjonsmekanismer for å fikse skadet DNA, og representerer krysskmomunikasjon mellom disse to kodene.
Den epitranscriptomiske koden og RNA -koden: epitranscriptomic modifikasjoner av RNA kan påvirke RNA -prosessering og stabilitet, og koble disse to kodene i genregulering.
Immunresponskoden og Inositol-fosfatkoden: Inositolfosfat signalering kan modulere immuncellefunksjon, noe som indikerer potensiell krysskommunikasjon mellom disse kodene i immunresponsen.
Signaltransduksjonskoden og signalintegrasjonskodene: Signaltransduksjonsveier involverer ofte flere signaler og svar, noe som antyder krysskommuniasjon og integrasjon mellom forskjellige signalkoder.
Transkripsjonsfaktorkoden og transkripsjonell reguleringskode: transkripsjonsfaktorer regulerer genuttrykk, og deres aktivitet påvirkes av forskjellige transkripsjonelle regulatoriske elementer, noe som viser krysskommunikasjon mellom disse kodene i genregulering.
Den ribosomale koden og proteinsekretorisk kode: Ribosomal aktivitet er essensiell for proteinsyntese og sekresjon, noe som indikerer krysskommuniasjon mellom disse kodene i proteinproduksjon og transport.
Bilde 6. Eks. på samarbeidende koder
 Følgende koder har ikke bare blitt karakterisert, men de krysskommuniserer:
Følgende koder har ikke bare blitt karakterisert, men de krysskommuniserer:
DNA-metyleringskoden og kromatinkoden: DNA-metylering kan påvirke kromatinstruktur og gentilgjengelighet. Metylgrupper tilsatt DNA kan rekruttere kromatinmodifiserende proteiner, noe som fører til endringer i kromatinorganisasjon og genuttrykk.
RNA-koden og proteinfoldingskoden: RNA-molekyler, så som ikke-kodende RNA og chaperon-assosierte RNA, kan påvirke proteinfolding og stabilitet. RNA -molekyler kan fungere som chaperoner, og hjelpe til med riktig proteinfolding og forhindre feilfolding.
Ubiquitin-koden og proteinfoldingskoden: Ubiquitin kan markere feilfoldede eller skadede proteiner for nedbrytning av proteasomet. Denne prosessen hjelper til med å opprettholde cellulært proteinkvalitetskontroll og forhindre akkumulering av toksiske proteinaggregater.
Signaltransduksjonskoden og cellesyklus sjekkpunktkode: Eksterne signaler mottatt under signaloverføring kan aktivere spesifikke sjekkpunkter i cellesyklusen, regulere celledeling og sikre nøyaktig DNA-replikasjon og kromosomsegregering.
DNA-reparasjonskoden og cellesyklus sjekkpunktkoden: DNA-skade kan aktivere cellesyklus sjekkpunkter, og stanse cellesyklusen for å gi tid til DNA-reparasjon før de fortsetter med celledeling.
Autofagi -koden og signaltransduksjonskoden: Visse signaloverføringsveier kan regulere autofagi som respons på næringstilgjengelighet eller cellulært stress.
DNA-reparasjonskoden og RNA-koden: Noen ikke-(protein) kodende RNA er involvert i DNA-reparasjonsprosesser og kan regulere ekspresjonen av gener som er involvert i DNA-reparasjon.
Kromatinkoden og RNA -koden: Kromatinmodifikasjoner kan påvirke transkripsjonen av RNA -molekyler, regulere genuttrykk og påvirke RNA -prosessering.
Forutsetning 1: Kompleksiteten og spesifisiteten ved cellulære koder, deres evne til å krysskommunisere og deres intrikate interaksjoner antyder et høyt nivå av koordinering og formål i utformingen av biologiske systemer.
Forutsetning 2: Fremveksten og funksjonaliteten til disse kodene kan ikke forklares tilstrekkelig med en trinnvis, gradvis og ikke-styrt evolusjonsprosess.
Konklusjon: Derfor innebærer eksistensen og egenskapene til disse cellulære kodene involvering av en intelligent designer i design og koordinering av komplekse flercellede organismer.
"Integrert kompleksitet: Argumentet for intelligent design i cellulære kommunikasjonssystemer" -Sammendrag via FB:
 Forutsetning 1: Cellulære koder fungerer i et svært sofistikert kommunikasjonsnettverk, og speiler intrikathetene til avanserte språk. Viktig for effektiv kommunikasjon er flere fullformede komponenter: en avsender som har sin opprinnelse, et overføringssystem som formidler meldingen, og en mottaker som tolker og handler på dem. Denne kommunikasjonsinfrastrukturen krever både maskinvare (de fysiske mekanismene som letter kommunikasjon) og programvare (den kodede informasjonen som blir videresendt). Hvert segment av dette systemet må være til stede og være omhyggelig kalibrert for å opprettholde integriteten og troskapen til budskapet, fra dets unnfangelse til dens kulminasjon.
Forutsetning 1: Cellulære koder fungerer i et svært sofistikert kommunikasjonsnettverk, og speiler intrikathetene til avanserte språk. Viktig for effektiv kommunikasjon er flere fullformede komponenter: en avsender som har sin opprinnelse, et overføringssystem som formidler meldingen, og en mottaker som tolker og handler på dem. Denne kommunikasjonsinfrastrukturen krever både maskinvare (de fysiske mekanismene som letter kommunikasjon) og programvare (den kodede informasjonen som blir videresendt). Hvert segment av dette systemet må være til stede og være omhyggelig kalibrert for å opprettholde integriteten og troskapen til budskapet, fra dets unnfangelse til dens kulminasjon.
Bilde 7. Eks. på kommuniasjon i celler
Forutsetning 2: I tillegg til denne kompleksiteten, viser disse cellulære kodene og språkene en evne til å "krysse" eller kommunisere på tvers av forskjellige veier eller systemer. I visse tilfeller kan en informasjonsstreng, avhengig av dens tolkning, formidle flere betydninger samtidig. Det tilsvarer en setning i menneskelige språk som bærer forskjellige betydninger basert på kontekst eller intonasjon. Slik mangefasettert kommunikasjon understreker videre dybden av presisjon og koordinering som ligger i disse systemene. Gitt disse lagdelte forviklingene, vil sannsynligvis mellomliggende eller delvis utviklede stadier være ikke-funksjonelle. En utviklende kode, avsendere uten harmoniserte mottakere, eller et umodent overføringssystem vil føre til feilkommunikasjon eller fullstendig kommunikasjonsfordeling. Dette kan sammenlignes med å prøve å utføre intrikat programvare på underutviklet maskinvare, eller et radioprogram med en vaklende sender eller mottaker. Følgelig ville disse delvise systemene, som mangler full funksjonalitet, ikke gi noen evolusjonær fordel. Dette scenariet utfordrer gjennomførbarheten av en stykkevis, ikke-styrt evolusjonsprosess for å lage et så integrert, multifunksjonelt kommunikasjonssystem.
Konklusjon: Gitt den lagdelte raffinementet av cellulær kommunikasjon, nødvendigheten av at hver komponent skal være fullt utviklet, og evnen til intrikat krysskommunikasjon og flere samtidige tolkninger, virker det mest plausibelt at en intelligent designer spilte en rolle i å konstruere dette koordinerte systemet innen komplekse flercellulære organismer.
Oversettelse via google oversetter og bilder ved Asbjørn E. Lund